Recombinant Drosophila melanogaster Neurogenic locus Notch protein (N), partial
-
中文名稱:黑腹果蠅N重組蛋白
-
貨號(hào):CSB-YP357053DLU
-
規(guī)格:
-
來源:Yeast
-
其他:
-
中文名稱:黑腹果蠅N重組蛋白
-
貨號(hào):CSB-EP357053DLU
-
規(guī)格:
-
來源:E.coli
-
其他:
-
中文名稱:黑腹果蠅N重組蛋白
-
貨號(hào):CSB-EP357053DLU-B
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:黑腹果蠅N重組蛋白
-
貨號(hào):CSB-BP357053DLU
-
規(guī)格:
-
來源:Baculovirus
-
其他:
-
中文名稱:黑腹果蠅N重組蛋白
-
貨號(hào):CSB-MP357053DLU
-
規(guī)格:
-
來源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:N; CG3936Neurogenic locus Notch protein [Cleaved into: Processed neurogenic locus Notch protein]
-
種屬:Drosophila melanogaster (Fruit fly)
-
蛋白長(zhǎng)度:Partial
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

靶點(diǎn)詳情
-
功能:Essential signaling protein which has a major role in many developmental processes. Functions as a receptor for membrane-bound ligands Delta and Serrate to regulate cell-fate determination. Upon ligand activation, and releasing from the cell membrane, the Notch intracellular domain (NICD) forms a transcriptional activator complex with Su(H) (Suppressor of hairless) and activates genes of the E(spl) complex. Regulates oogenesis, the differentiation of the ectoderm and the development of the central and peripheral nervous system, eye, wing disk, muscles and segmental appendages such as antennae and legs, through lateral inhibition or induction. Regulates neuroblast self-renewal, identity and proliferation through the regulation of bHLH-O proteins; in larval brains, involved in the maintenance of type II neuroblast self-renewal and identity by suppressing erm expression together with pnt; might also regulate dpn expression through the activation of the transcriptional regulator Su(H).
-
基因功能參考文獻(xiàn):
- Proper assembly of multicellular mechanosensory organs requires a double-negative circuit involving miRNA-mediated suppression of a Notch repressor to assign non-neuronal cell fate. PMID: 29196461
- Study shows that the trans- and cis- interactions between the ligand Ser and the Notch receptor regulate the cell number and size of the imaginal ring in drosophila. PMID: 29773559
- Strat is required to localize Rab8 at the TGN, and rab8 phenocopies strat We propose that Strat and Rab8 act at the exit of the Golgi apparatus to regulate the sorting and the polarized distribution of Notch, Delta and Spdo. PMID: 29967125
- Delta-Notch dependent boundary formation in the Drosophila large intestine PMID: 28950873
- The authors found that Dl requires ubi for its full function, but can also signal in two ubi-independent modes, one dependent and one independent of Neur. Their analysis suggests that one important role of Dl protein ubiquitination by Mib1 is their release from cis-inhibitory interactions with Notch, enabling them to trans-activate Notch on adjacent cells. PMID: 28960177
- AKAP200 is a novel tissue specific posttranslational regulator of Notch, maintaining high Notch protein levels and thus promoting Notch signaling. PMID: 29309414
- In the nervous system the subperineurial glia (SPG) are unique in using both the endocycle and endomitosis to grow. In the brain, the majority of SPG initially endocycle, then switch to endomitosis during larval development. The Notch signaling pathway and the String Cdc25 phosphatase are crucial for the endocycle versus endomitosis choice. PMID: 29440220
- During ovarian germline stem cell niche formation, the status of Notch signaling in the cell can be reprogrammed. This is controlled via steroid-induced miR-125, which targets a negative regulator of Notch signaling, Tom. PMID: 29361571
- Notch-Abl axonal signaling requires proteolytic cleavage events that initiate canonical Notch signaling. Some Notch protein is tyrosine phosphorylated, this form of the protein is selectively associated with Disabled and Trio, and relevant tyrosines are essential for Notch-dependent axon patterning but not for canonical Notch-dependent regulation of cell fate. PMID: 29343637
- Data show that the wave speed control and salt and pepper pattern formation required distinct threshold levels of Notch signaling activity. PMID: 27535937
- Regulation of NOTCH signaling by RAB7 and RAB8 requires carboxyl methylation by ICMT. PMID: 29051265
- Grh, AbdA, and Exd function together at multiple motifs on the apoptotic enhancer. Exd-Grh-Notch work with different Hox genes through region specific enhancers to pattern respective segments of larval central nervous system. PMID: 29023471
- Inscuteable maintains type I neuroblast lineage identity via Numb/Notch signaling in the Drosophila larval brain PMID: 28325554
- and ecdysone signaling pathways synergistically regulate Cut expression for proper DV boundary formation in the wing disc PMID: 27117286
- biochemical analysis shows that A2BP1 is part of the Suppressor of Hairless [Su(H)] complex in the presence and absence of Notch PMID: 28174239
- Nerfin-1 represses Notch activity in medulla neurons and prevents them from dedifferentiation. PMID: 28242614
- reciprocal relationship between Notch signalling and cell cycle progression acts like a developmental clock, providing a delimited window of time during which cells decide their fate, ensuring efficient and orderly bristle patterning. PMID: 27226324
- This work not only elucidates a key mechanism of Notch-mediated maintenance of type II NB self-renewal and identity, but also reveals a novel function of Erm. PMID: 27151950
- To identify structural principles underlying polyQ tracts in disordered regulatory domains, study analyzed deep evolution of metazoan Notch polyQ tracts. Results suggest that intrinsically disordered interference arrays featuring carboxamide and polyQ enrichment may constitute coupled proteodynamic modulators of solenoids. PMID: 28319202
- During antennal development, expression of the selector genes Lim1 and Dll was sharply segregated. Notch signaling induces actomyosin-dependent apical constriction and epithelial fold. Disruption of Notch signaling or the actomyosin network reduces apical constriction and epithelial fold. PMID: 28708823
- our data indicate that the MLF/DnaJ-1-dependent increase in Lz level allows the repression of Notch expression and signaling to prevent aberrant blood cell development. Thus our findings establish a functional link between MLF and the co-chaperone DnaJ-1 to control RUNX transcription factor activity and Notch signaling during blood cell development in vivo. PMID: 28742844
- The conserved MAPK site in E(spl)-M8, an effector of Drosophila notch signaling, controls repressor activity during eye development. PMID: 27428327
- Notch signaling directly regulates ban expression at the transcriptional level to impact cell growth. PMID: 28520736
- Loss of Usp5 results in upregulation of Notch signaling and downregulation of RTK signaling by EGF receptor (EGFR) and Sevenless (Sev), leading to impaired photoreceptor development. PMID: 28140449
- These data provide foundational information for future studies investigating the mechanisms of how O-glycosylation regulates Notch activity. PMID: 27268051
- N activity is required to promote dpn transcription; only in R7 photoreceptor precursors does the removal of Ttk coincide with high N activity; and only in this cell does Dpn expression result PMID: 27427987
- lysosomal-associated functions regulated by the TFEB-V-ATPase axis might play a conserved role in shaping cell fate. PMID: 26727288
- the monosaccharide O-glucose and terminal dixylose of O-glucose-linked saccharides have distinct activities in Notch trafficking, although a loss of these activities is compensated for by the presence of monosaccharide O-fucose. PMID: 27129198
- Rabex-5 plays an important role in Drosophila hematopoiesis and might serve as an axis coordinating Ras and Notch signaling in the lymph gland. PMID: 26567216
- Reducing levels of old intermediate neural progenitor (INP) temporal transcription factor Eyeless/Pax6 allows Notch signaling to promote dedifferentiation of INP progeny into ectopic INPs, creating a proliferative mass of ectopic progenitors in the brain. PMID: 26585279
- Drosophila adult muscle precursors display homing behavior to muscle niche and the niche-driven Insulin-Notch-dMyc cascade plays a key role in setting the activated state of adult muscle precursors. PMID: 26650355
- Data suggest that gliolectin is a positive regulator of Notch receptor signalling during wing development. PMID: 26505251
- a Notch/Su(H)/E(spl)-HLH cascade specifically controls daughter, but not progenitor proliferation PMID: 27070787
- Data show that Notch protein functions solely by the canonical pathway in regulating physiological plasticity. PMID: 26986723
- endocycle entry in anterior follicle cells than those in the posterior, identified that the insulin-PI3K pathway participates in the precise M/E switch, and suggested Nejire as a cofactor of Notch signaling during oogenesis PMID: 26205122
- Notch protein regulates Dys expression during Drosophila leg development. PMID: 25329825
- NICD signaling is not sufficient to explain Nfull-induced Long-term memory enhancement. PMID: 25791355
- We further show that Notch in ORNs acts by both canonical cleavage-dependent and non-canonical cleavage-independent pathways. The Notch ligand Delta (Dl) in PNs switches the balance between the pathways. PMID: 26011623
- Significantly, the JNK pathway is responsible for the majority of the phenotypes and transcriptional changes downstream of Notch-Src synergy. PMID: 26222204
- The role of the Hippo pathway effectors Yorkie and Scalloped in mediating and facilitating Notch signaling-mediated lineage specification in the lymph gland was studied. PMID: 26151599
- we show that the binding of the fly RAM domain of the NICD to CSL does not affect interactions of the corepressor Hairless with CSL PMID: 25650119
- Myoblast cytonemes mediate Wg signaling from the wing imaginal disc and Delta-Notch signaling to the air sac primordium. PMID: 25951303
- By altering the ratios between Yki, Sd and Vg, Notch pathway activation restricts the effects of Yki mediated transcription. PMID: 25157415
- A novel role of Notch signaling in controlling germline stem cell-niche adhesion in response to aging. PMID: 25521289
- Notch activation leads to robust increase in H3K56 acetylation. PMID: 26069324
- A branched pathway downstream of Notch in which Bowl functions to direct hub cell assembly in parallel to Tj downregulation, is reported. PMID: 26092848
- Notch directly regulates the cell morphogenesis genes Reck, talin and trio in adult muscle progenitor PMID: 25217625
- Data indicate that Notch signaling pathway components are expressed in adult malpighian tubules. PMID: 25527105
- study reveals that Chip is a novel interacting partner of Notch PMID: 25597954
- Notch signaling activates the expression of extramacrochaetae (emc), which encodes a helix-loop-helix (HLH) transcription factor. Emc, in turn, then forms a biochemical complex with Da. PMID: 25977368
顯示更多
收起更多
-
亞細(xì)胞定位:Cell membrane; Single-pass type I membrane protein. Endosome. Note=Transported to early endosomes by O-fut1 (PubMed:17329366).; [Processed neurogenic locus Notch protein]: Nucleus. Note=Upon activation and S3 cleavage, it is released from the cell membrane and enters into the nucleus in conjunction with Su(H).
-
蛋白家族:NOTCH family
-
數(shù)據(jù)庫(kù)鏈接:
KEGG: dme:Dmel_CG3936
STRING: 7227.FBpp0070483
Most popular with customers
-

Recombinant Human CD226 antigen (CD226), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
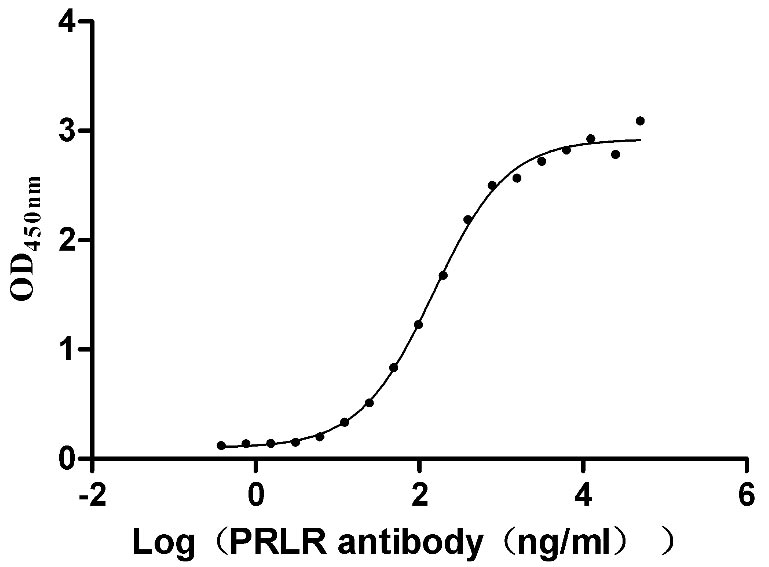
Recombinant Human Prolactin receptor (PRLR), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
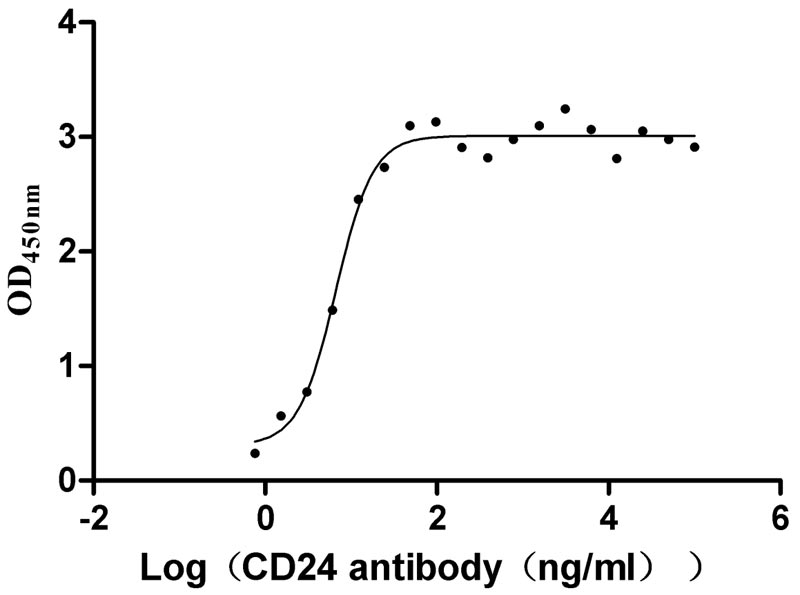
Recombinant Human Signal transducer CD24 (CD24)-Nanoparticle (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
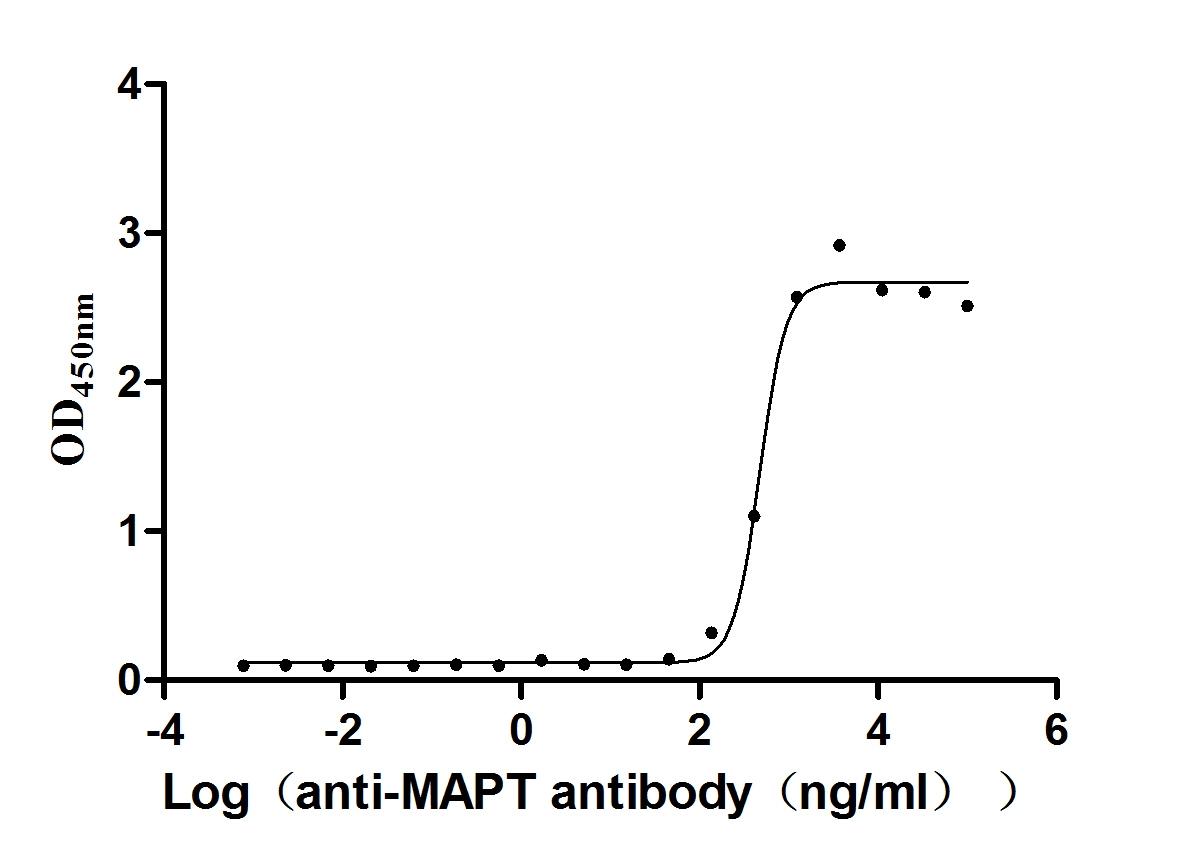
Recombinant Mouse Microtubule-associated protein tau (Mapt) (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
-AC1.jpg)
Recombinant Human Microtubule-associated protein tau (MAPT) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
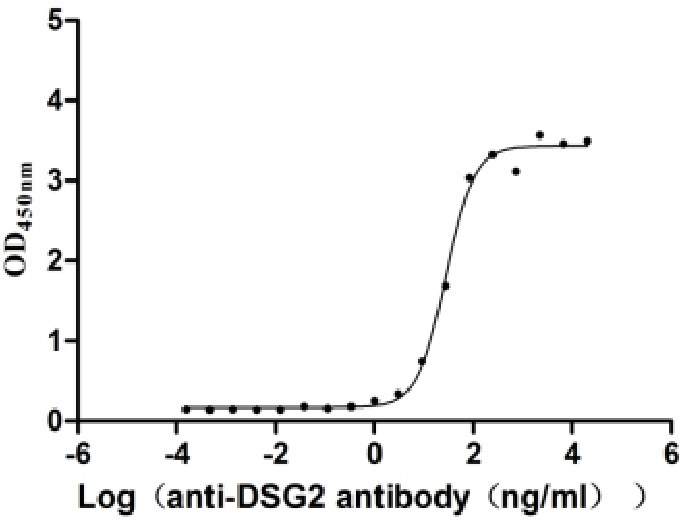
Recombinant Human Desmoglein-2 (DSG2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
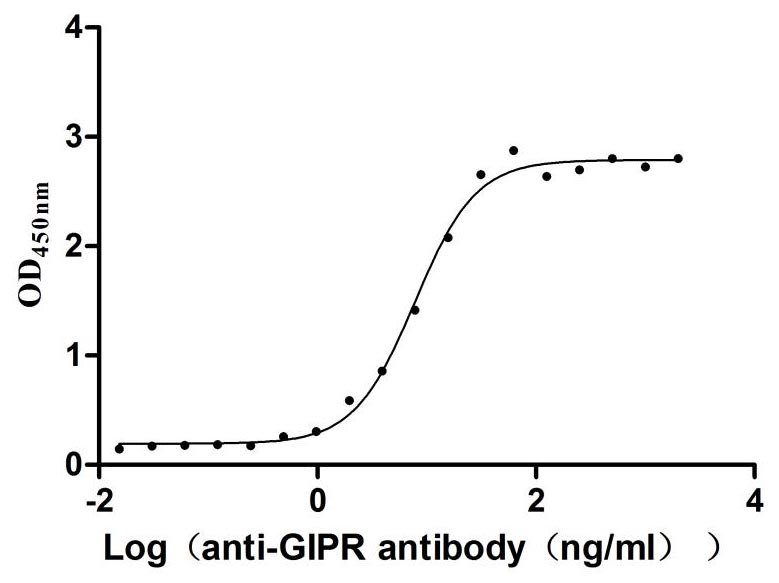
Recombinant Rat Gastric inhibitory polypeptide receptor (Gipr), partial (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
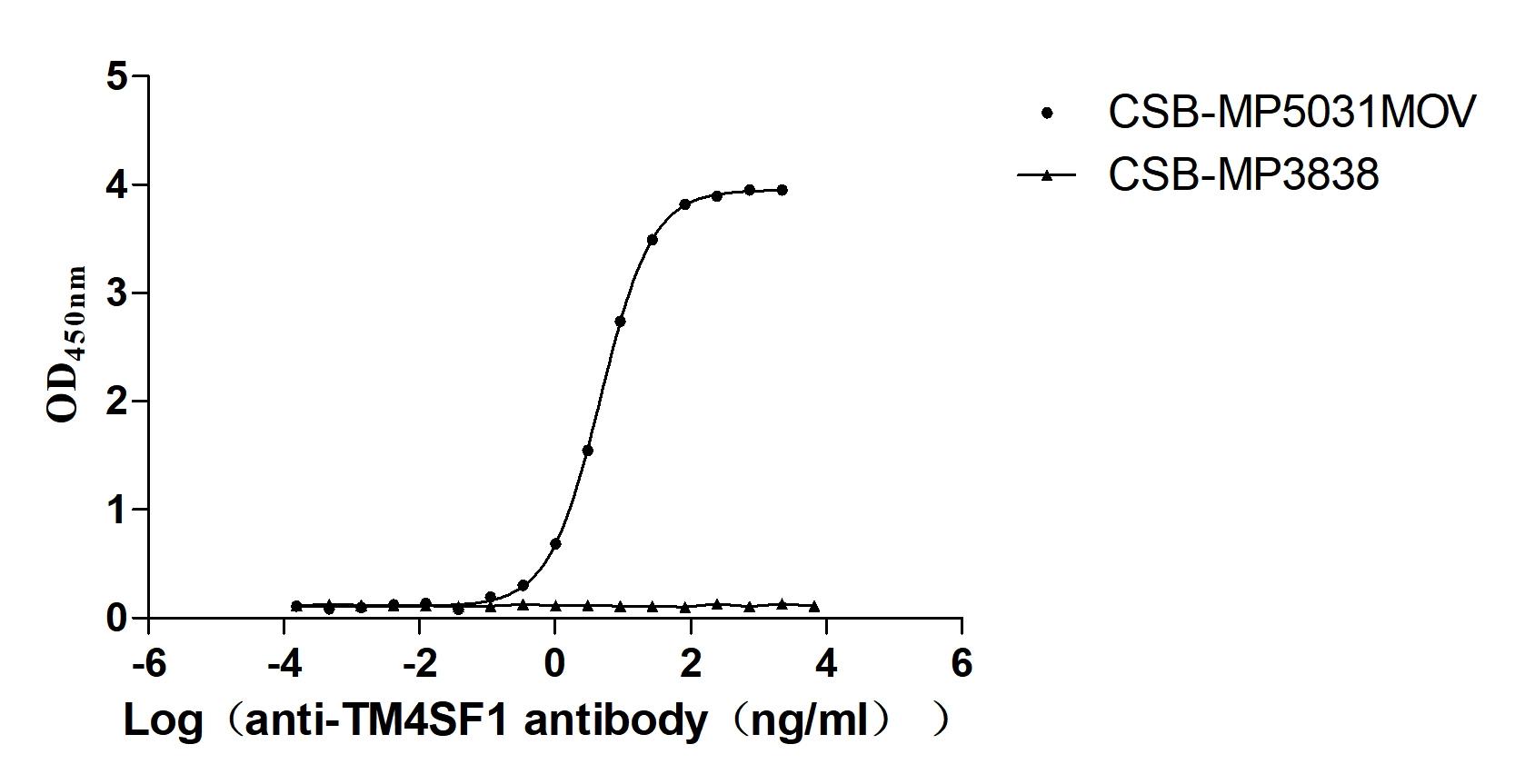
Recombinant Macaca fascicularis Transmembrane 4 L6 family member 1 (TM4SF1)-VLPs (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)









