Recombinant Drosophila melanogaster Armadillo segment polarity protein (arm), partial
-
中文名稱:Recombinant Drosophila melanogaster Armadillo segment polarity protein(arm) ,partial
-
貨號(hào):CSB-YP325615DLU
-
規(guī)格:
-
來源:Yeast
-
其他:
-
中文名稱:Recombinant Drosophila melanogaster Armadillo segment polarity protein(arm) ,partial
-
貨號(hào):CSB-EP325615DLU
-
規(guī)格:
-
來源:E.coli
-
其他:
-
中文名稱:Recombinant Drosophila melanogaster Armadillo segment polarity protein(arm) ,partial
-
貨號(hào):CSB-EP325615DLU-B
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Drosophila melanogaster Armadillo segment polarity protein(arm) ,partial
-
貨號(hào):CSB-BP325615DLU
-
規(guī)格:
-
來源:Baculovirus
-
其他:
-
中文名稱:Recombinant Drosophila melanogaster Armadillo segment polarity protein(arm) ,partial
-
貨號(hào):CSB-MP325615DLU
-
規(guī)格:
-
來源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:arm
-
Uniprot No.:
-
別名:arm; CG11579Armadillo segment polarity protein
-
種屬:Drosophila melanogaster (Fruit fly)
-
蛋白長(zhǎng)度:Partial
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

靶點(diǎn)詳情
-
功能:Isoform neural may associate with CadN and participate in the transmission of developmental information. Can associate with alpha-catenin. Isoform cytoplasmic accumulates through wg signaling; arm function in wg signal transduction is required early in development for determination of neuroblast fate. Arm and Abl proteins function cooperatively at adherens junctions in both the CNS and epidermis.
-
基因功能參考文獻(xiàn):
- Study showed that the highly evolutionary conserved Y654 beta-cat-D665-E-cad major interaction site of the adherens junctions complex acts as primary mechanosensor. The site activates the beta-cat pathway through its mechanical opening in vivo that enables phosphorylation by Src42A kinase. PMID: 30024850
- Manipulating Axin or APC2 levels had no effect on beta-catenin destruction complex activity when Wnt signals were absent, but, surprisingly, had opposite effects on the destruction complex when Wnt signals were present. Elevating Axin made the complex more resistant to inactivation, while elevating APC2 levels enhanced inactivation. PMID: 29641560
- here we examine the interactions between Msk and the Wg pathway in regulation of the AMP pool size. We find that a myoblast-specific reduction of Msk results in the absence of Vg expression and a complete loss of the Wg pathway readout beta-catenin/Armadillo (Arm). Moreover, msk RNA interference knockdown abolishes expression of the Wg target Ladybird (Lbe) in leg disc myoblasts. PMID: 28249984
- An absolute requirement for Armadillo for activation and repression of Wingless protein target genes PMID: 28369070
- The data suggests that arm-Gal4 has detrimental effects on Drosophila development and lifespan that are directly dependent upon parental inheritance PMID: 26505429
- dissociation of tyrosine-phosphorylated Arm from DE-cadherin allows dynamic actin to reorganize, leading to ring canal expansion and cell shape changes during the course of oogenesis PMID: 25803041
- Armless may promote Wg signaling by rescuing Armadillo from proteolytic degradation. PMID: 25369031
- Reduced Klp64D function is suppressed by activated Arm. PMID: 25063455
- Arm activity strongly suppresses supernumerary neuroblasts induced by overexpression of klu. PMID: 24257623
- Results implicate a previously unrecognized function for miR-310/13 in dampening the activity of Arm in early somatic and germline progenitor cells, whereby inappropriate/sustained activation of Arm-mediated signaling or cell adhesion may impact normal differentiation in the Drosophila male gonad. PMID: 23821034
- Arm is found at the plasma membrane of all epithelial cells, as part of the cadherincatenin complex PMID: 22359584
- Binding of Arm to DE-cadherin is weaker in polarizing cells than in polarized cells. PMID: 22159415
- Data show that Hipk dually regulates both Wingless and Hedgehog signaling by impeding the function of the E3 ubiquitin ligase complex, thereby inhibiting Armadillo ubiquitination and subsequent degradation. PMID: 21628596
- In the current model of Wnt signalling, a complex assembled around Axin and Apc allows GSK3 (Shaggy) to phosphorylate Armadillo and target it for degradation. PMID: 21389052
- the Hippo pathway restricts Wnt/beta-Catenin signaling by promoting an interaction between TAZ and DVL in the cytoplasm PMID: 20412773
- Data show that in arm mutants, even a modest reduction in the basolateral component lgl leads to a full apical domain expansion or lgl phenotype. PMID: 19737741
- proper polarity in the late embryo involves the asymmetric distribution and phosphorylation of Armadillo at the membrane, and interference with this Arm phosphorylation leads to polarity defects PMID: 17183721
- Data show that wingless (wg) represses transcription of the dpp gene in the ventral leg disc, and that this repression requires a tri-partite complex of the WG mediators armadillo and dTCF, and the co-repressor Brinker. PMID: 17206277
- Control of beta-catenin phosphorylation/degradation by a dual-kinase mechanism PMID: 11955436
- MyoVI is required for border cell migration where it stabilizes E-cadherin and Arm. PMID: 12134162
- The Drosophila clock protein Timeless is a member of the Arm/HEAT family. PMID: 12372263
- we find a strong correlation between progression through mitosis and a reduction in Armadillo levels.We conclude that this phenomenon may reduce the efficacy of Wingless signalling and/or intercellular adhesion during cell division. PMID: 12559488
- elucidation of signaling in nucleus PMID: 12646868
- Results show that a vital regulatory step in Wingless signaling is Zeste white 3-mediated Armadillo phosphorylation. PMID: 14966281
- Tws is required for the stabilization of Armadillo/beta-catenin in response to Wg/Wnt signaling. PMID: 14973271
- armadillo/beta-catenin has a nuclear function PMID: 15024404
- Pygo and Lgs function in targeting Armadillo/beta-catenin to the nucleus, thus ensuring its availability to TCF during Wnt signalling. PMID: 15208637
- Armadillo/beta-catenin-dependent Wnt signalling is required for the polarisation of epidermal cells during dorsal closure in Drosophila PMID: 15226252
- Data show that Drosophila Hyrax and its human ortholog, Parafibromin, are required for nuclear transduction of the Wnt/Wg signal and bind directly to the C-terminal region of beta-catenin/Armadillo. PMID: 16630820
- The activity of armadillo is regulated by Notch synergizing with axin in Drosophila. PMID: 16881048
- binding per se of beta-catenin by adenomatous polyposis coli does not require phosphorylation by GSK-3beta PMID: 17968317
- Wg, beta-catenin, and decapentaplegic regulate the division of embryonic PGCs in a stage-specific manner PMID: 18291628
- Data show that Drosophila Naked cuticle (Nkd) engages the nuclear import adaptor Importin-alpha3 to antagonize Wnt/beta-catenin signaling. PMID: 18423435
- Mbt activation destabilizes Drosophila Armadillo with DE-cadherin resulting in a decrease in DE-cadherin-mediated adhesiActivation of Mbt leads to destabilization of Drosophila Armadillo with DE-cadherin resulting in decreasing cadherin-mediated adhesion. PMID: 18636970
- In vivo evidence for a complex non-linear elationship between Armadillo levels, subcellular distribution and Wingless signalling. PMID: 18682750
- Bone morphogenetic protein pathway components can repress Wg target gene expression by influencing the binding of Arm and dTcf. PMID: 19065265
- Using both in vitro and in vivo assays, Hipk was found to promote the stabilization of Arm. PMID: 19088090
- Arm is a major regulator of cadherin-mediated adhesion and is cleaved by caspases in embryo extracts and study provides evidence that the caspase-3 homolog drICE cleaves Arm in vitro and in vivo. PMID: 19232093
- Data suggest that DE-cadherin/armadillo form part of the mechanism that control branching and trajectory of axon tracts in the larval brain. PMID: 19520071
- Data have identified PR55 alpha as the regulatory subunit of PP2A that controls beta-catenin phosphorylation and degradation. PR55 alpha, but not the catalytic subunit, PP2Ac, directly interacts with beta-catenin. PMID: 19556239
- Notch can promote the degradation of activated forms of Armadillo and may buffer cells against fluctuations in Wnt signalling activity. PMID: 19668359
- differential requirements of contact residues in Axin for interactions with GSK3beta or beta-catenin. PMID: 19850033
顯示更多
收起更多
-
亞細(xì)胞定位:Cytoplasm. Cell membrane; Peripheral membrane protein; Cytoplasmic side. Cell junction, adherens junction. Note=Inner surface of cell membrane and adherens junction.
-
蛋白家族:Beta-catenin family
-
組織特異性:Isoform cytoplasmic accumulates at low levels in axons, at high levels in specific cells along the CNS midline and in leg and eye imaginal disks. Isoform neural accumulates in the axon tracts of the CNS. Both isoforms accumulate in the peripheral nervous
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
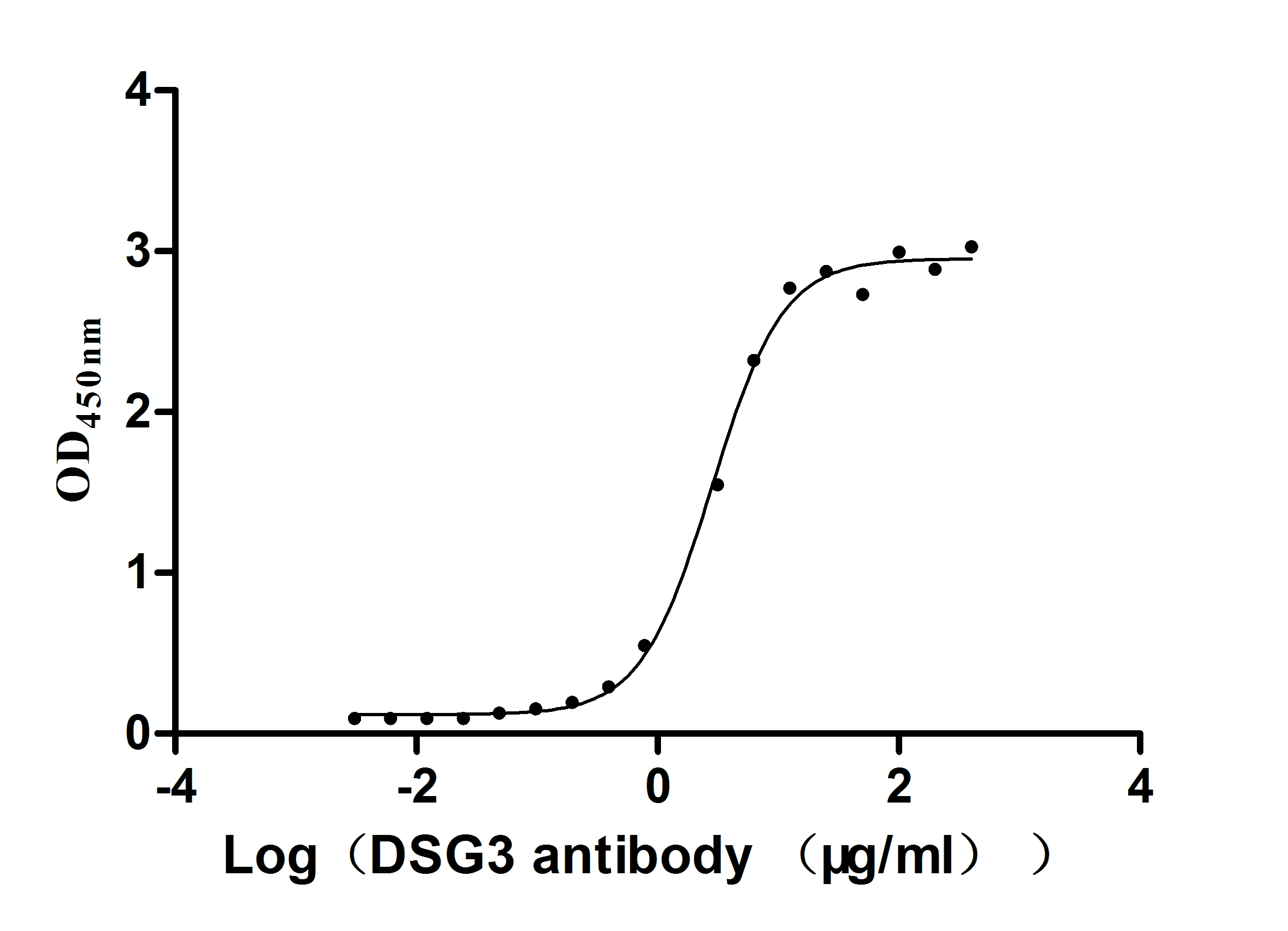
Recombinant Mouse Desmoglein-3 (Dsg3), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
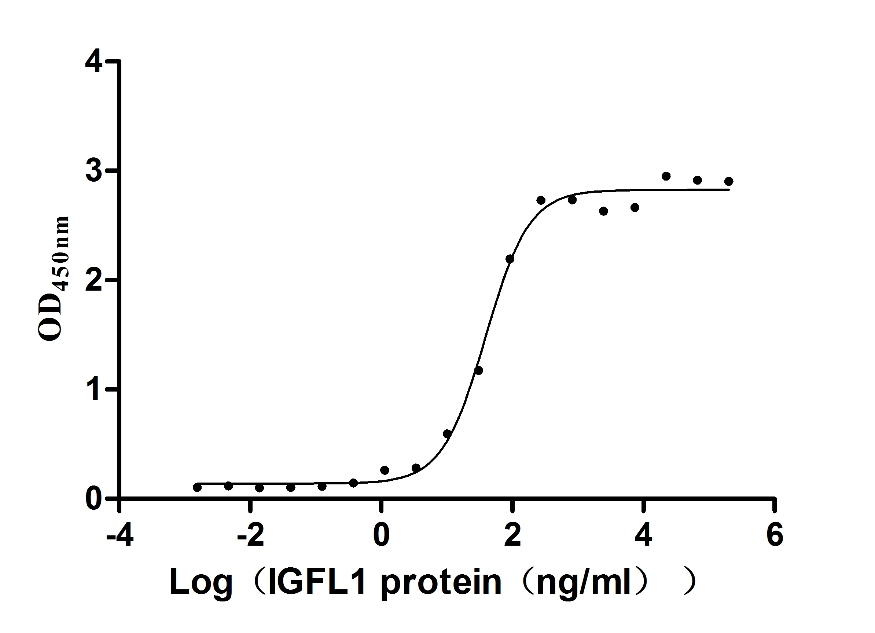
Recombinant Human Insulin growth factor-like family member 1 (IGFL1) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
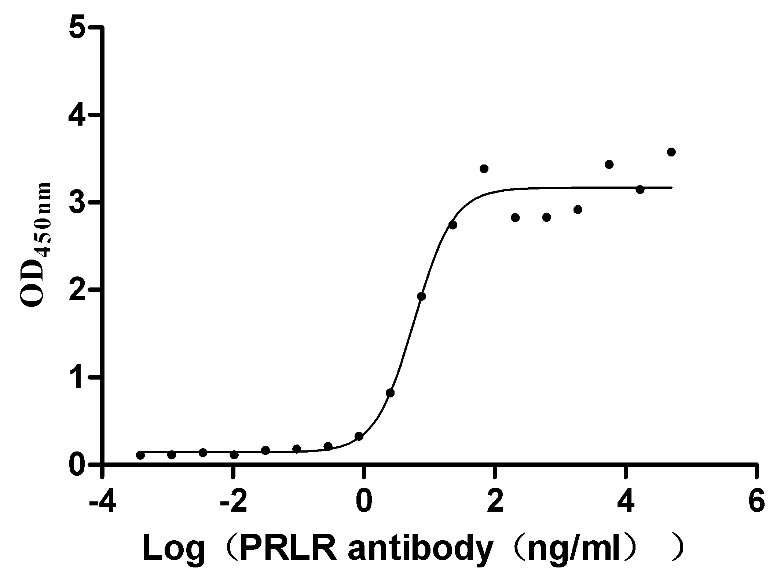
Recombinant Mouse Prolactin receptor (Prlr), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
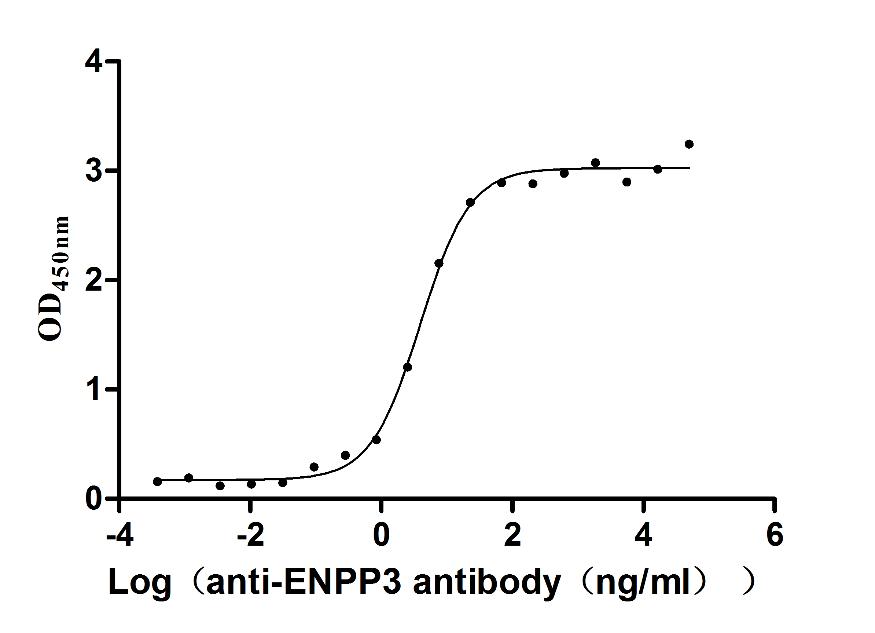
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
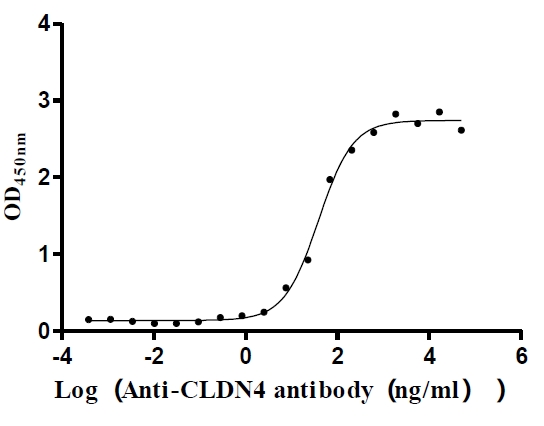
Recombinant Human Claudin-4 (CLDN4)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
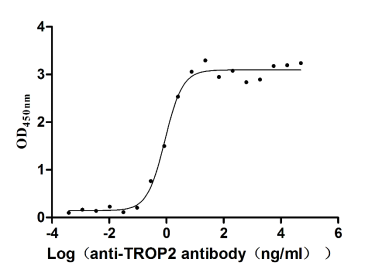
Recombinant Human Tumor-associated calcium signal transducer 2 (TACSTD2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
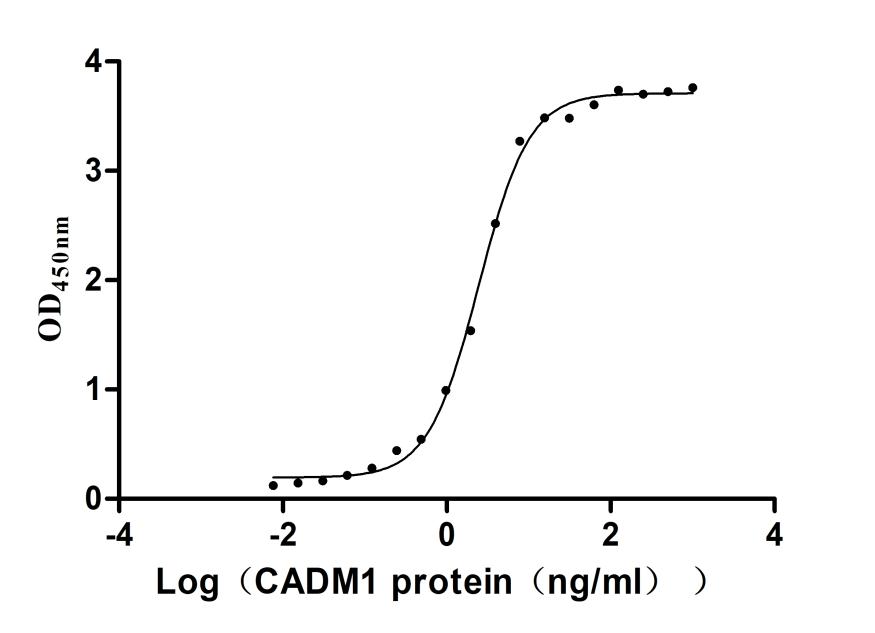
Recombinant Human Cytotoxic and regulatory T-cell molecule (CRTAM), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
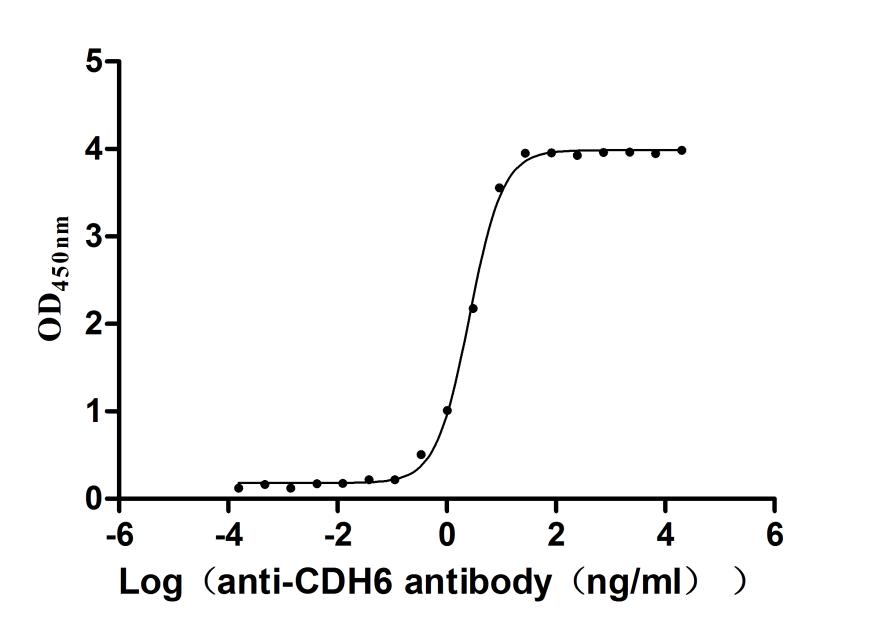
Recombinant Human Cadherin-6(CDH6),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









