Recombinant Drosophila melanogaster Target of rapamycin (Tor), partial
-
中文名稱:黑腹果蠅Tor重組蛋白
-
貨號(hào):CSB-YP892859DLU
-
規(guī)格:
-
來(lái)源:Yeast
-
其他:
-
中文名稱:黑腹果蠅Tor重組蛋白
-
貨號(hào):CSB-EP892859DLU
-
規(guī)格:
-
來(lái)源:E.coli
-
其他:
-
中文名稱:黑腹果蠅Tor重組蛋白
-
貨號(hào):CSB-EP892859DLU-B
-
規(guī)格:
-
來(lái)源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:黑腹果蠅Tor重組蛋白
-
貨號(hào):CSB-BP892859DLU
-
規(guī)格:
-
來(lái)源:Baculovirus
-
其他:
-
中文名稱:黑腹果蠅Tor重組蛋白
-
貨號(hào):CSB-MP892859DLU
-
規(guī)格:
-
來(lái)源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:Tor
-
Uniprot No.:
-
別名:Tor; CG5092; Serine/threonine-protein kinase Tor; EC 2.7.11.1; Target of rapamycin
-
種屬:Drosophila melanogaster (Fruit fly)
-
蛋白長(zhǎng)度:Partial
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

靶點(diǎn)詳情
-
功能:Promotes cell and tissue growth, maintains tissue homeostatis and controls responses to environmental stress and aging. Regulates growth during animal development by coupling growth factor signaling to nutrient availability. Central regulators of autophagy. May be involved in atg1 phosphorylation. May also be involved, directly or indirectly, in the control of neuronal function. Phosphorylates S6K/p70S6K, in vitro. May regulate the activity of S6K. Overexpression inhibits growth and reduces cell size. Affects the timing of neuronal cell differentiation. Hyperactivation of the signaling leads to accelerated differentiation, whereas inhibition of the signaling retards differentiation. Thus, in addition to controlling growth of the cell in which it resides, it can also influence growth of distant cells and organs during development via a humoral mechanism. As part of the TORC1 complex regulates energy homeostasis and promotes certain aspects of larval growth by negatively regulating REPTOR. REPTOR functions downstream of TORC1 to regulate the expression of stress response genes in response to TORC1 inhibition resulting from nutrient deprivation. When TORC1 activity is high it phosphorylates REPTOR which inhibits its recruitment into the nucleus and antagonizes their function. This function is essential under normal feeding conditions to promote TORC1-dependent growth during larval development and, in adults and larvae to prevent the REPTOR-dependent expression of nutrient stress response genes. In short, during development, it primarily controls growth, whereas in the adult, where there is relatively little growth, it controls aging and other aspects of nutrient-related physiology. Rag GTPases act as activators of TORC1 in response to amino acid signals.
-
基因功能參考文獻(xiàn):
- TORC1 is selectively activated in the second mitotic wave of the developing Drosophila eye. TORC1 is regulated by Hedgehog and E2F1 pathway. PMID: 28829944
- TORC1 activity in the wing disc is patchy. TORC1 activity is high in cells at the G1/S transition due to CycD/Cdk4. PMID: 28829945
- We show in vivo that autophagy is impaired in dGBA-deficient fly brains. In response, mechanistic target of rapamycin (mTOR) activity is downregulated in dGBA-deficient flies and rapamycin ameliorates the lifespan, locomotor, and oxidative stress phenotypes. dGBA knock-out flies also display an upregulation of the Drosophila ortholog of mammalian TFEB, Mitf, a response that is unable to overcome the autophagy block. Toget PMID: 27852774
- To characterize natural genetic variation in the IIS/TOR pathway, we used >250 recombinant inbred lines (RILs) derived from a multiparental mapping population, the Drosophila Synthetic Population Resource, to map transcript-level QTL of genes encoding 52 core IIS/TOR components in three different nutritional environments PMID: 28592498
- This study demonstrated that the absence of an engulfment receptor leads to a pronounced accumulation of dead neurons in the brain of the fruit fly and Rescued by TORC1 Activation. PMID: 26985028
- evidence points to an ancient module comprising Mitf, v-ATPase and TORC1 that serves as a dynamic modulator of metabolism for cellular homeostasis. PMID: 26092939
- Data show that genetic reduction in TOR Complex 1 (TORC1) signalling improves the impaired motor performance phenotype of Friedreich's ataxia (FRDA) model flies. PMID: 26158631
- Loss of TORC2 disrupted the nuclear localization of Myc, and inhibited Myc-dependent transcription. PMID: 25999153
- We show that changing the proportion of t(6)A-modified tRNAi (Met), by expression of an un-modifiable tRNAi (Met) or changing the levels of Tcs3, regulate target of rapamycin (TOR) kinase activity and influences cell and animal growth in vivo. PMID: 26063805
- lowered dietary amino acids promote longevity via TOR PMID: 24861087
- nutrient sensing through the Insulin/IGF and TOR pathways couples cuticle pigmentation of both male and female Drosophila with their nutritional status during metamorphosis. PMID: 24133012
- our observation established that Spargel/dPGC-1 is indeed a terminal effector in the insulin-TOR pathway operating below TOR, S6K, Tsc, and FoxO PMID: 23934892
- Tor and InR are required cell-autonomously for the proliferation of precursors for both somatic niches and germline stem cells. PMID: 24026119
- DREF is required for growth downstream of TOR, but not insulin/PI3K signaling. PMID: 22960233
- Target of rapamycin (Tor) mutants did show a phenotype that mimicked WT starvation-induced PCD, indicating an insulin independent regulation of PCD via Tor signaling. PMID: 22240900
- Loss of postsynaptic TOR disrupts a retrograde compensatory enhancement in neurotransmitter release that is normally triggered by a reduction in postsynaptic glutamate receptor activity. PMID: 22500638
- Cellular detoxification pathways, increased autophagy and altered protein synthesis have all been implicated in increased lifespan from reduced IIS/TOR activity, with the role of defence against oxidative stress unresolved PMID: 20849947
- we review the contribution of the model organism Drosophila in the understanding of TOR signaling and the various biological processes it modulates that may impact on aging PMID: 21130151
- TORC1-S6K-RPS6 (as a fusion protein)signaling axis is regulated by many subcellular components, including the Class I vesicle coat (COPI), the spliceosome, the proteasome, the nuclear pore, and the translation initiation machinery. PMID: 21239477
- These results uncover specific TOR functions in the control of stem cells versus their differentiating progeny, and reveal parallels between Drosophila and mammalian follicle growth. PMID: 20504961
- analysis of Dendritic tiling through TOR signalling PMID: 20010972
- Rheb functions in the insulin signaling pathway upstream of TOR. PMID: 12766776
- Loss of Tsc1 and Tsc2 or ectopic activation of Rheb results in constitutive activation of TOR and renders S6K activity resistant to amino acid starvation. PMID: 12771962
- results suggest that dTOR and dRictor have a shared positive role in the phosphorylation of the hydrophobic motif site of dAkt/dPKB PMID: 15718470
- tap42 functions independently of target of rapamycin to regulate cell division and survival in Drosophila PMID: 15802506
- TOR-mediated cell-cycle activation causes neurodegeneration in a Drosophila tauopathy model, identifying TOR and the cell cycle as potential therapeutic targets in tauopathies and Alzheimer Disease. PMID: 16461276
- Endocytosis acts both as an effector function downstream of TOR and as a physiologically relevant regulator of TOR signaling. PMID: 16785324
- Here, they show that reducing the function of Drosophila TOR results in decreased lipid stores and glucose levels. PMID: 16890541
- Results show that mildly increasing systemic Rheb-TOR-S6K signaling sensitizes the whole organism to oxidative stress and promotes senescence of locomotor activity with age. PMID: 17038544
- Rheb-TOR signaling controls S2 cell growth by promoting ribosome production and protein synthesis, not by direct effects on the import of amino acids or glucose. The effect of insulin signaling upon TOR activity varies by cellular type and context. PMID: 17371599
- Results show that the effects of TOR on growth and metabolism was mediated by Myc. PMID: 18177722
- a function of Rag GTPases in TORC1 activation in response to amino acid signals PMID: 18604198
- The Tor pathway couples nutrition and developmental timing in Drosophila. PMID: 18854141
- Studies show the increasingly prominent links between TOR signaling and aging in invertebrates. PMID: 19539012
- up-regulation of dEif4e is sufficient to recapitulate the effects of high dTOR or insulin signaling in cardiac aging in Drosophila. PMID: 19594484
- Data indicate that TOR induces cell death by suppressing autophagy and provide direct genetic evidence that autophagy alleviates cell death in several common types of neurodegenerative disease. PMID: 19720874
顯示更多
收起更多
-
蛋白家族:PI3/PI4-kinase family
-
數(shù)據(jù)庫(kù)鏈接:
Most popular with customers
-
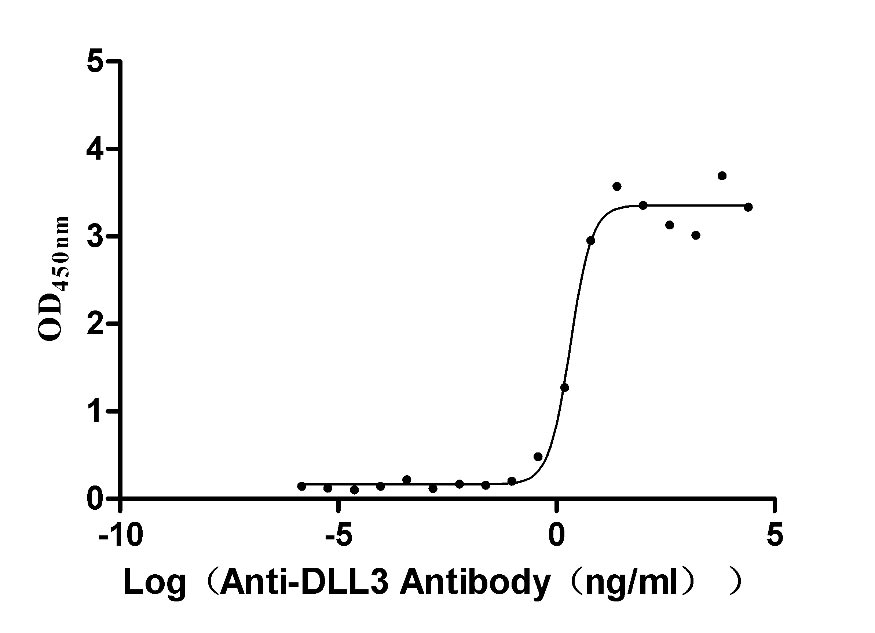
Recombinant Macaca fascicularis Delta-like protein 3 (DLL3), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
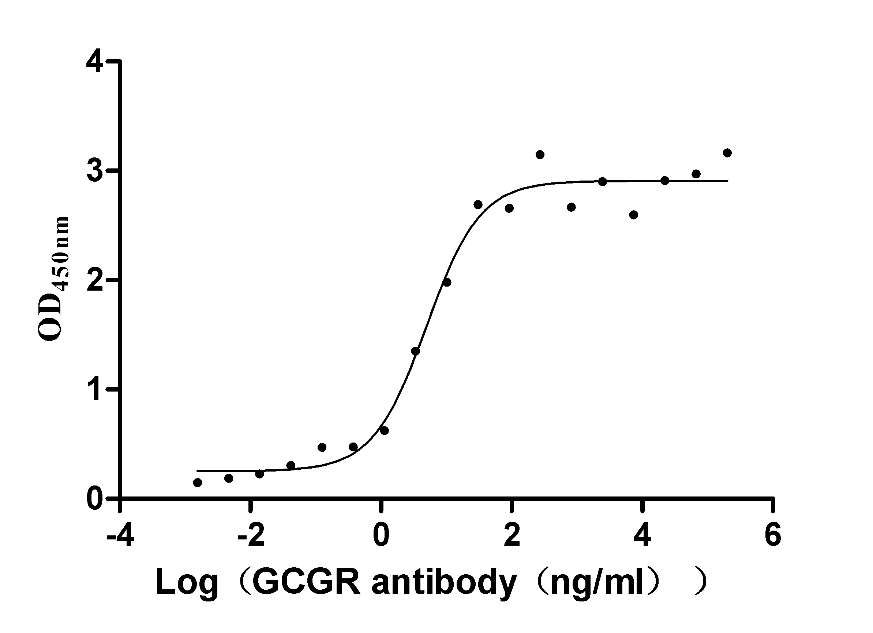
Recombinant Human Glucagon receptor (GCGR), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Human Angiopoietin-2 (ANGPT2) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
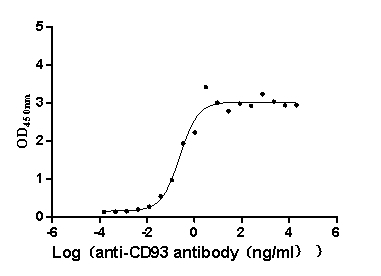
Recombinant Macaca fascicularis CD93 molecule (CD93), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
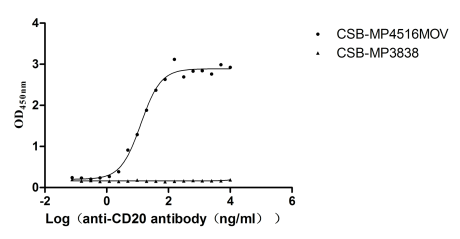
Recombinant Macaca fascicularis Membrane spanning 4-domains A1 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Human Urokinase-type plasminogen activator(PLAU) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









